| This article appears courtesy of the New Mexico Museum of Natural History and Science. "Maryland's Dinosaur" web site, "Dinosaur Fund" and Dr. Kranz, are not affiliated with or endorsed by the New Mexico Museum of Natural History and Science (NMMNHS) or the NMMNHS Bulletins. These pages may not be reproduced or redistributed in any form. |
Lucas, S.G., Kirkland, J.I. and Estep, J.W., eds., 1998, Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin No. 14. 225-228 pp. |
New Mexico Museum of Natural History, 1801 Mountain Road, Albuquerque, NM 87104
ABSTRACT: Acrocanthosaurus, from the Trinity Group of Oklahoma and Texas, remains the only adequately known large theropod from the Lower Cretaceous of North America, though large theropod bones have been recovered from the Cloverly, Arundel, and Cedar Mountain Formations. All the known clearly diagnostic elements (except some teeth) from these units pertain to taxa that are not congeneric with Acrocanthosaurus. The Arundel, Cloverly, and Cedar Mountain formations appear to be, at least in part, approximately contemporaneous with Trinity Group sediments. As a result, Acrocanthosaurus was not the only large theropod inhabiting North America during the Early Cretaceous.
In contrast with small theropod taxa (e.g., Deinonychus [Ostrom, 1969] and Microvenator [Ostrom, 1970]), large theropods in the Lower Cretaceous of North America remain poorly known. This situation contrasts sharply with those of other dinosaur-bearing formations on the continent; for example, in the Upper Jurassic Morrison Formation, large theropods (e.g., Allosaurus and Ceratosaurus) are represented by more numerous and more complete specimens than contemporaneous small theropods (e.g., Ornitholestes, Coelurus, and EIaphrosaurus).
To date, the only large theropod from the Lower Cretaceous of North America known from relatively complete, diagnostic material is Acrocanthosaurus atokensis. The holotype and paratype specimens of this taxon are fragmentary skeletons from the Antlers Formation of Oklahoma (Stovall and Langston, 1950); the taxon has recently been redescribed based on a more complete skeleton (Harris, 1997, 1998). The comparative, if not actual, completeness of material of Acrocanthosaurus has led several authors (e.g., Pittman, 1989; DeCourten, 1991) to attribute isolated large theropod elements from Lower Cretaceous sediments in North America to Acrocanthosaurus or a similar taxon without identifying in those elements any autapomorphies of the taxon. The tacit assumption is that Acrocanthosaurus is the only large theropod present in the Lower Cretaceous of North America. However, material from other large theropods does exist, and can be shown to belong to taxa other than Acrocanthosaurus.
Abbreviations used: CEU = College of Eastern Utah, Price; FWMSH = Ft. Worth Museum of Science and History, Ft. Worth; OMNH = Oklahoma Museum of Natural History; SMU = Southern Methodist University, Dallas; USNM = United States National Museum, Smithsonian Institution, Washington, D.C.; UUVP = University of Utah, Salt Lake City; YPM = Yale Peabody Museum, New Haven
The first Early Cretaceous large theropod material described from North America was from the Arundel Formation (Potomac Group) of Maryland. Marsh (1888) based the species Allosaurus medius on teeth, an astragalus, and a phalanx. Lull (1911) attributed vertebrae to the taxon; Lull et al. (1911) specified four vertebrae (a ?mid- or ?caudal dorsal, a proximal caudal, and two distal caudals) and two phalanges, all from the same locality, as the original material. Lull et al. (1911) referred all the material, except a single tooth, attributed to the taxon by Marsh (1888) to the ornithopod Dryosaurus. (The material was shown to be ornithomimosaurian by Gilmore [1920,1921], and the phalanges were referred to the genus Archaeornithomimus by Russell [1972; q.v. Kranz, 1989.] Having removed all elements except the tooth from A. medius, Lull et al. (1911) then attributed a caudal dorsal, a proximal caudal, and two distal caudal vertebrae as well as two pedal phalanges to the taxon. Gilmore (1920,1921) noted that the two distal caudals belong to an ornithomimosaurian and again asserted that only the tooth (USNM 4972) could be typical of A. medius. Gilmore (1921) removed the tooth from the genus Allosaurus and referred to it as ?Dryptosaurus medius. The tooth lacks an adequate description, and its status is indeterminate.
Lull (1911) described a second large Arundel theropod, Creosaurus potens, on the basis of a vertebral centrum (USNM 3049) larger than those assigned to A. medius. Lull et al. (1911) described it as a caudal dorsal vertebra, but it was shown by Gilmore (1921) to be a proximal caudal. Gilmore (1921) noted that the genus Creosaurus was congeneric with Allosaurus, and that the isolated Arundel centrum was more similar to the Late Cretaceous theropod Dryptosaurus aquilunguis (Cope, 1866; Marsh, 1877; Carpenter et al., 1997), so he called the Arundel form ?Dryptosaurus potens. The Arundel centrum lacks any medial constriction and possesses a single, strong ventral keel (Gilmore, 1921). In contrast, caudals of Acrocanthosaurus have arched ventral margins, and where keeled, have a low, double keel (Harris, 1997, 1998). The Arundel centrum lacks the rudimentary pleurocoelus fossae present on the proximal caudal centra of Acrocanthosaurus as well (Fig. 1). Thus, there is a large theropod present in the North American Early Cretaceous that is distinct from Acrocanthosaurus. It remains unknown whether or not this caudal vertebra belongs to the same taxon as the tooth of ?Dryptosaurus medius. Weishampel and Young (1996: 132) consider all the nonornithomimosaur theropod material from the Arundel Formation indeterminate, but it is at least clear that the caudal vertebra is not congeneric with Acrocanthosaurus.
Large, isolated theropod teeth from the Arundel are reportedly (T. Lipka, personal communication, 1997) virtually identical to those of Acrocanthosaurus described by Harris (1997,1998), so this genus may be present in the Arundel fauna, as well.
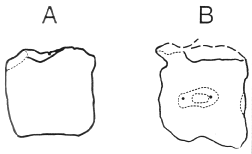
FIGURE 1. A, Caudal vertebral centrum, USNM 3049, referred to ?Dryptosaurus potens, from the Arundel Formation of Maryland (after Gilmore, 1921). Centrum length = 14 cm. B, Eighth caudal vertebral centrum, SMU 74646 FWMSH 93B-9 1-12, of Acrocanthosaurus (from Harris, 1997). Centrum length = 13.5 cm. Not to scale.
Ostrom (1970: 69-70, fig. 6) described a dorsal vertebra (YPM 5285) belonging to an "indeterminate theropod" from the Cloverly Formation of Montana. This specimen is markedly different from dorsals of Acrocanthosaurus described by Harris (1997,1998). The vertebra is not opisthocoelous, as are the cranial dorsals of Acrocanthosaurus. While the Cloverly vertebra's transverse processes do have a dorsocaudal angulation, it is not as strong as in Acrocanthosaurus. Its centrum lacks pleurocoels, and the transverse process is not invaded by the large, deep fossae seen in Acrocanthosaurus (Harris, 1997, 1998). The pre- and postzygapophyses of the Cloverly form are small and simple, unlike the large, arched facets of Acrocanthosaurus. The neural spine is not elongate, and expands craniocaudally towards its summit, both unlike Acrocanthosaurus (Fig. 2). Overall, the Cloverly vertebra appears to have originated with a theropod more primitive than Acrocanthosaurus. ,As noted by Ostrom (1970), the Cloverly dorsal cannot be compared with the proximal caudal vertebra of ?Dryptosaurus potens from the Arundel to assess congenericity. If it represents a distinct taxon, then there is a second large, non-Acrocanthosaurus theropod present in the Early Cretaceous of North America.
DeCourten (1991: fig. 5) illustrates a tooth (UUVP 904) from the Long Walk Quarry in the Cedar Mountain Formation (Ruby Ranch Member per Kirkland et al., 1997) of Utah attributed to Acrocanthosaurus or a similar taxon. Kirkland and Parrish (1995) and Kirkland et al. (1997) state their belief that the tooth morphology is distinct from Acrocanthosaurus. Examination of this specimen indicates that it differs from the Acrocanthosaurus tooth described by Harris (1997, 1998) in possessing approximate 1 denticle per mm, as opposed to 2 per mm in the Trinity taxon's tooth. However, precise measurements could not be taken, and examination of the specimen was impaired by incomplete preparation between the denticles. Furthermore, without description of complete tooth-bearing skull elements for Acrocanthosaurus, it is unclear if there is differentiation between teeth at various positions in the mouth: the denticle size difference between the Cedar Mountain and Trinity teeth may simply be an artifact of heterodonty.
Kirkland et al. (1997) note the presence of an large theropod ilium, also from the Long Walk Quarry. The association of the ilium in the same quarry as the teeth may indicate that they originated from the same taxon, if not the same individual. As no ilium of Acrocanthosaurus has yet been described, it is impossible to determine if the specimen represents the Trinity taxon, the taxon represented in the quarry by the coarsely-serrated teeth, or a different large theropod. If the ilium and the teeth belong to the same taxon, then it remains distinct from Acrocanthosaurus. The table provided by Kirkland et al. (1997, p. 79, table 2) states that "cf. Acrocanthosaurus sp." is present in the Ruby Ranch fauna; this is in reference to an isolated tooth (CEU 5107) with very fine serrations from the Cedar Mountain Formation near the Cleveland-Lloyd Dinosaur Quarry (J. Kirkland, personal comm., 1998). The table also lists a "new large theropod" in the Ruby Ranch fauna, in reference to the taxon represented by the coarsely-serrated teeth.
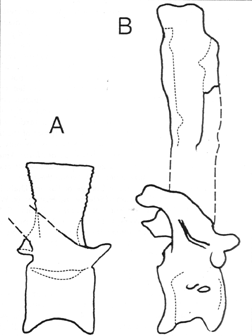
FIGURE 2. A, Dorsal vertebra, YPM 5285, of a large, unidentified theropod from the Cloverly Formation of Montana (after Ostrom, 1970). Centrum length = 9.5 cm. B, Ninth dorsal vertebra, SMU 74646/FWMSH 93B-9 A1-2-14 (centrum) and 2A-3 (neural spine; height based on OMNH 8-O-S9) of Acrocanthosaurus (from Harris, 1997). Centrum length = 13.5 cm. Drawings not to scale.
Acrocanthosaurus is known from the Twin Mountain Formation of Texas (Harris, 1997,1998) and the Antlers Formation of Oklahoma (Stovall and Langston, 1950). The Aptian-Albian boundary, dated to 112+/-0.l Ma by Obradovich (1994) and Gradstein et al. (1994, 1995), can be identified, based in part on ammonite biostratigraphy (Young, 1986), in the Glen Rose Limestone that overlies the Twin Mountains Formation in Texas (Jacobs et al., 1991; Winkler et al., 1995). A late Aptian-early Albian age for the group, and a late Aptian age for the Twin Mountains Formation in particular, is supported by 13C correlation data (Rennison, 1996; see also Jacobs and Winkler, in press).
The age of the Arundel Formation has been listed as lateAptian or early Albian (Kranz, 1989,1996); this age, for at least a portion of the dinosaur-bearing strata, is supported by palynostratigraphy (Robbins, 1991; Doyle, 1992). Thus, the large theropod represented by the caudal called ?Dryptosaurus potens, as well as the tooth of ?Dryptosaurus medius may be approximately contemporaneous with Acrocanthosaurus.
Ostrom (1970) used vertebrate biostratigraphy to date the Cloverly Formation to the late Aptian-early Albian. Cloverly and Trinity Group sediments share the dinosaur genera Deinonychus (Cifelli et al., 1997b) and Tenontosaurus (Winkler et al., 1997), the chelonian Naomichelys (Ostrom, 1970), and several related mammals (Jacobs et al., 1991; Cifelli, 1993); however, some of the shared taxa differ at the species level (Cifelli et al., 1997b; Winkler et al., 1997), and Naomichelys is of dubious biostratigraphic utility due to its long temporal distribution (Cifelli et al., 1997a). Comparison of the mammalian faunas shows that the Cloverly fauna may be contemporaneous or slightly younger than the Trinity (Jacobs et al., 1991; Jacobs and Winkler, in press). Fission-track dating of zircons from volcanic tuffs in the Cloverly have yielded ages as old as the Valanginian (~ 137-132 Ma: Gradstein et al., 1995) at its base to late Aptian-middle Albian and younger at the top (Chen and Lubin, 1997). Ostrom (1970) finds many Cloverly taxa to have a relatively wide stratigraphic distribution, and the range of the poorly-known large theropod represented by the dorsal vertebra is unknown: it could be somewhat older, contemporaneous, or slightly younger than Acrocanthosaurus.
The Cedar Mountain Formation records a large portion of the Early Cretaceous and at least part of it, the Ruby Ranch Member, appears to be correlative with the Trinity Group based on faunal similarities (Kirkland, 1996; Kirkland et al., 1997; Cifelli et al., 1997a). Although the absolute age of the Ruby Ranch Member is not yet available for comparison with the well-constrained age of the Trinity fauna (Jacobs and Winkler, in press), the faunal similarities (Kirkland et al., 1997) likely indicate a similar age.
Although diagnostic material of Acrocanthosaurus atokensis has not yet been identified from sediments outside of the Trinity Group, large theropods are found in other Lower Cretaceous rocks in North America. Described material, including a proximal caudal vertebral centrum from the Arundel Formation of Maryland (?Dryptosaurus potens) and a caudal dorsal vertebra from the Cloverly Formation of Montana, are clearly distinct from Acrocanthosaurus. A tooth from the Ruby Ranch Member of the Cedar Mountain Formation of Utah also appears distinct. Therefore, there is at least one, and possibly as many as three, large theropods in addition to Acrocanthosaurus present in the Lower Cretaceous of North America.
ACKNOWLEDGMENTS
Thanks are due to Louis Jacobs and Dale Winkler (Southern Methodist University) who suggested and oversaw this research as part of my Master of Science thesis. Peter Kranz and Tom Lipka provided information on both old and new material from the Arundel. Jim Kirkland (Dinamation International) generously allowed me to study specimens from the Cedar Mountain Formation in his care. Louis Jacobs, Dale Winkler, Thomas Holtz, and Spencer Lucas provided helpful reviews of this manuscript.
This work was completed in partial fulfillment of the requirements for a Masters Degree at Southern Methodist University.
REFERENCES
Carpenter, K., Russell, D., Baird, D., and Denton, R. 1997. Redescription of the holotype of Dryptosaurus aquilunguis (Dinosauria: Theropoda) from the Upper Cretaceous of New Jersey. Journal of Vertebrate Paleontology 17: 561-573.
Chen, Z.-Q. and Lubin, S. 1997. A fission-track study of the terrigenous sedimentary sequences of the Morrison and Cloverly Formations in the northeastern Bighorn Basin, Wyoming. The Mountain Geologist 34: 51-62.
Cifelli, R.L. 1993. Early Cretaceous mammal from North America and the evolution of marsupial dental characters. Proceedings of the National Academy of Sciences 90: 9413-9416.
Cifelli, R.L., Kirkland, J.I., Well, A., Deino, A.L., and Kowallis, B.J. 1997a. High-precision 40Ar/39Ar geochronology and the advent of North America's Late Cretaceous terrestrial fauna. Proceedings of the National Academy of Sciences 94: 11163-11167.
Cifelli, R.L., Gardner, J.D., Nydam, R.L., and Brinkman, D.L. 1997b. Additions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous), southeastern Oklahoma. Oklahoma Geology Notes 57: 124-131.
Cope, E.D. 1866. Remarks on the remains of a gigantic extinct dinosaur from the Cretaceous Greensand of New Jersey. Proceedings of the Academy of Natural Sciences of Philadelphia 18: 275-279.
DeCourten, F. 1991. The Long Waik quarry and tracksite: unveiling the mysterious Early Cretaceous of the Dinosaur Triangle region, pp. 19-25 in Averett, W.R. (ed.) Guidebook for Dinosaur quarries and tracksites tour, western Colorado and eastern Utah. Grand Junction: Grand Junction Geological Society.
Doyle, J.A. 1992. Revised palynological correlations of the lower Potomac Group (USA) and the Cocobeach sequence of Gabon (Barremian-Aptian). Cretaceous Research 13:337-349.
Gilmore, C.W. 1920. Osteology of the carnivorous Dinosauria in the United States National Museum with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus. United States National Museum Bulletin 110: l-154.
Gilmore, C.W. 1921. The fauna of the Arundel Formation of Maryland. Proceedings of the Umted States National Museum 59: 581-594.
Gradstein, FM., Agterberg, F.P., Ogg, J.G., Hardenbol, J., van Veen, P., Thierry, J., and Huang, Z. 1994. A Mesozoic time scale. Journal of Geophysical Research 99: 24,051-24,074.
Gradstein, FM., Agterberg, F.P.. Ogg, J.G., Hardenbol, J., van Veen, P., Thierry, J., and Huang, Z. 1995. A Triassic, Jurassic, and Cretaceous time scale, pp. 95-126 in Berggen, W.A., Kent, D.V., and Aubry, M.-P. (eds.) Geochronology, time scales, and global stratigraphic correlation. SEPM Special Publication 54.
Harris, J.D. 1997. A reanalysis of Acrocanthosaunis atokensis, its phylogenetic relationships, and paleobiogeographic implications, based on a new specimen from Texas. Unpublished MS Thesis, Southern Methodist University, 204 pp.
Harris, J.D., 1998. A reanalysis of Acrocanthosaurus Atokensis, its phylogenetic relationships, and paleobiogeographic implications, based on a new specimen from Texas. New Mexico Museum of Natural History Bulletin 13: l-75.
Jacobs, L.L. and Winkler, D.A. In press. Mammals, archosaurs, and the Early to Late Cretaceous transition in north-central Texas, in Tomida, Y., Flynn, L.J., and Jacobs, L.L. (eds.) Papers on vertebrate paleontology and chronology in honor of Everett H. Lindsay. National Science Museum, Tokyo.
Jacobs, L.L., Winkler, D.A., and Murry, P.A. 1991. On the age and correlation of Trinity mammals, Early Cretaceous of Texas, USA. Newsletter on Stratigraphy 24: 35-43.
Kirkland, J.I. 1996. Biogeography of western North America's mid-Cretaceous dinosaur faunas: losing European ties and the first great Asian-North American interchange. Journal of Vertebrate Paleontology 16 (suppl. 3): 45A.
Kirkland, J.I. and Parrish, J.M. 1995. Theropod teeth from the lower and middle Cretaceous of Utah. Journal of Vertebrate Paleontology 15 (suppl. 3): 39A.
Kirkland, J.I., Britt, B., Burge, D.L., Carpenter, K., Cifelli, R., DeCourten, F., Eaton, J., Hasiotis, S., and Lawton, T. 1997. Lower to middle Cretaceous dinosaur faunas of the central Colorado Plateau: a key to understanding 35 million years of tectonics, sedimentology, evolution, and biogeography. Brigham Young University Geology Studies 42 (part II): 69-103.
Kranz, P.M. 1989. Dinosaurs in Maryland. Maryland Geological Survey Department of Natural Resources Educational Series 6: l-34.
Kranz, P.M. 1996. Notes on the sedimentary iron ores of Maryland and their dinosaurian fauna. Studies in Maryland Geology Special Publication of the Maryland Geological Survey 3: 87-115.
Lull, R.S. 1911. The Reptilia of the Arundel Formation. Maryland Geological Survey, Lower Cretaceous: 174-178.
Lull, R.S., Clark, W.B., and Berry, E.W. 1911. Systematic paleontology of the Lower Cretaceous deposits of Maryland: Vertebrata. Maryland Geological Survey, Lower Cretaceous: 183-211.
Marsh, O.C. 1877. Notice of a new and gigantic dinosaur. American Journal of Science (ser. 3) 114: 87-88.
Marsh, O.C. 1888. Notice of a new genus of Sauropoda and other new dinosaurs from the Potomac Formation. American Journal of Science (ser. 3) 35: 89-94.
Obradovich, J.D. 1994. A Cretaceous time scale, pp. 379-396 in Caldwell, W.G.E. and Kauffman, E.G. (eds.) Evolution of the Western Interior Basin. Geological Association of Canada Special Paper 39.
Ostrom, J.H. 1970. Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin Area, Wyoming and Montana. Peabody Museum of Natural History Bulletin 35: l-234.
Pittman, J.G. 1989. Stratigraphy, lithology, depositional environment, and track type of dinosaur track-bearing beds of the Gulf coastal plain, pp. 135-153 in Gillette, D.D. and Lockiey, M.G. (eds.) Dinosaur Tracks and Traces. Cambridge University Press, Cambridge.
Rennison, C.J. 1996. The stable carbon isotope record derived from mid-Cretaceous terrestrial plant fossils from north-central Texas. Unpublished MS thesis, Southern Methodist University, 110 pp.
Robbins, E.I. 1991. Age of Early Cretaceous palynomorphs in the Muirkirk Clay Pit fossil locality (Prince Georges County, MD). U.S. Geological Survey Open File Report 91-613: l-7.
Russell, D.A. 1972. Ostrich dinosaurs from the Late Cretaceous of western Canada. Canadian Journal of Earth Sciences 9: 375-402.
Stovall, J.W. and Langston, W. 1950. Acrocanthosaurus atokensis, a new genus and species of Lower Cretaceous Theropoda from Oklahoma. American Midland Naturalist 43: 696-728.
Weishampel, D.B. and Young, L. 1996. Dinosaurs of the East Coast. Baltimore: Johns Hopkins University Press, 275 pp.
Winkler, D.A., Jacobs, L.L., and Murry, P.A. 1997. A new species of Tenontosaurus (Dinosauria: Ornithopoda) from the Early Cretaceous of Texas. Journal of Vertebrate Paleontology 17: 330-348.
Winkler, D.A., Jacobs, L.L., Lee, Y.-N., and Murry, P.A. 1995. Sea level fluctuation and terrestrial faunal change in north-central Texas, pp. 175-177 in Sun, A. and Wang Y. (eds.) Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers. China Ocean Press,Beijing.
Young, K. 1986. Cretaceous, marine inundations of the San Marcos Platform, Texas. Cretaceous Research 7: 117-140.