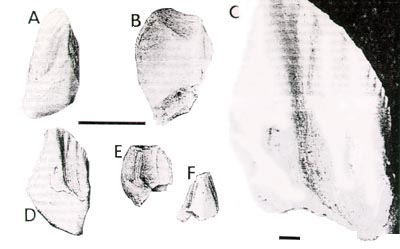
Scale for A, B, E, F = 1cm, for C=1 mm.
| This article appears courtesy of the New Mexico Museum of Natural History and Science. "Maryland's Dinosaur" web site, "Dinosaur Fund" and Dr. Kranz, are not affiliated with or endorsed by the New Mexico Museum of Natural History and Science (NMMNHS) or the NMMNHS Bulletins. These pages may not be reproduced or redistributed in any form. |
Lucas, S.G., Kirkland, J.I. and Estep, J.W., eds., 1998, Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin No. 14. 297-302 pp. |
ABSTRACT: Among recent discoveries of new neoceratopsian material in North America are teeth from the Cedar Mountain Formation of Utah (Albian-Cenomanian boundary) and the Arundel Formation of Maryland (late Aptian). These teeth are determined to belong to Neoceratopsia on the basis of gross morphology. A character suite, which includes a bulbous, convex non-enameled crown surface, deep indentations, a well-developed cingulum, and secondary ridges that end within the cingulum, distinguishes neoceratopsian from similar ornithopod teeth. These characters, when studied in combination with other morphological traits, can be confidently used for identification of isolated neoceratopsian teeth. The early and continent-wide appearance of Neoceratopsia in North America necessitates a reevaluation of the biogeography of the clade.
The uppermost Cedar Mountain Formation of eastern Utah has been dated at 98.39 +/- 0.07 years (right at the Albian-Cenomanian boundary: Obradovich, 1993) through the use of radiometric methods (Cifelli et al., 1997). Although previously thought to be unfossilferous (Stokes, 1952; Young, 1960), these strata have recently yielded a diverse terrestrial fauna including the earliest occurring hadrosaurid (Kirkland, this volume) and some of the earliest known ceratopsian material (Cifelli et al., 1997; in press). The latter consists of isolated teeth, determined here to belong to Neoceratopsia.
Neoceratopsian teeth have now been identified from the Arundel Clay facies of the Potomac Formation at the Muirkirk fossil site in central Maryland (the stratigraphic and depositional aspects of the Arundel are covered in Lipka, this volume). The first Arundel tooth specimen was collected in 1990 by Peter Kranz in Prince Georges County, Maryland (USNM 337977; Fig. 2). Although initially thought to belong to Dryosauridae or Tenontosaurus (Kranz, 1996) one of us [M. B.-S.] recognized that it might possibly belong to a ceratopsian. A second Maryland tooth, USNM 497708 (Fig. 3), was collected in 1997 by one of us (T. L.). This tooth is unworn, unlike the first one, and provides additional information for taxon determination.
Morphological comparisons are made with teeth from a variety of ornithopod and ceratopsian taxa, with the resulting conclusion that the Cedar Mountain and Arundel teeth are most likely neoceratopsian. Ornithopod and ceratopsian teeth vary a great deal among their included taxa. In neoceratopsians, teeth also vary among individuals and even within tooth rows (Maryanska and Osmolska, 1975). Although it is difficult to identify characters that are apomorphic for the clade, this study has successfully identified several characters that can diagnose confidently neoceratopsian teeth. These include a bulbous, convex shape of the non-enameled or less- enameled side of the crown, the presence of indentations or "pockets" that are deepest closest to the root, a well-developed cingulum enclosing the indentations, a primary ridge distally offset in maxillary teeth and mesially in mandibular teeth, secondary ridges that terminate within the indentations, and vertical wear facets. Not all neoceratopsian teeth possess all of the above characters, and some of them are present on ornithopod teeth. It is therefore a combination of traits that must be used to determine the taxonomic identity of new specimens.
Biogeographic implications of the presence of Neoceratopsia in the Early Cretaceous of Utah and Maryland include several possibilities. The presence of neoceratopsians on the east coast of the continent provides evidence for a pre-Aptian or early Aptian fauna1 migration from Eurasia, either across from Asia via the Bering land bridge (although this bridge was most likely not in place until the middle Cretaceous [Cifelli et al., 1971]) or westward across Europe. If ancestors of the Arundel neoceratopsians arrived in western North America from Asia, then a dispersal event is also required from the western part of the continent eastward before the inland seaway divided the land and made passage impossible. The other possibility is that of a North American origin for the clade, instead of the widely recognized Asian origin (Dodson and Currie, 1990).
Abbreviations: AMNH, American Museum of Natural History, New York; USNM, United States National Museum of the Smithsonian Institution, Washington DC; GPIT, Geologica Paleontologica Institut Tubingen, Germany; YPM, Yale Peabody Museum, New Haven, Connecticut; OU, Oklahoma University, Norman, Oklahoma; UUVP, Utah University Vertebrate Paleontology Collections, Salt Lake City, Utah; MNA, Museum of Northern Arizona, Flagstaff, Arizona; OMNH, Oklahoma Museum of Natural History, Norman, Oklahoma; MOR, Museum of the Rockies, Bozeman, Montana.
MNA v9023 represents a tooth from an immature animal or from the very front or back of a tooth row. It is much smaller than the other Cedar Mountain tooth and exhibits less development of shape, primary and secondary ridges, cingulum and indentations. Dimensions of the teeth are listed in Table 1.
Buccally, the crowns of the Cedar Mountain (Fig. 1) and Arundel (Figs. 2-3) teeth have an ivy-leaf-shape, close to that of some ornithopods (Tenontosaurus sp., OU 8-0-S3; Hypsilophodon: Norman, 1984) and nearly all neoceratopsians, especially Leptoceratops (Brown, 1914; Gilmore, 1939). Many ornithopod teeth have crowns much taller in comparison to mesiodistal width, including teeth of some members of Heterodontosauridae (Weishampel and Witmer, 1990), Telmatosaurus (Weishampel et.al., 1993), Iguanodon (Norman, 1984), and Dryosaurus (GPIT 1713/21-29). Hypsilophodon teeth are wider than neoceratopsian teeth (Norman, 1984), and hadrosaurid teeth are extremely derived and lozenge-shaped (Norman, 1984).
| USNM 337977 |
USNM 497708 |
MNA v9023 |
USNM 28179 |
|
| Crown height | 12.72 | 10.58 | 8.34e | 13.84 |
| Max. mesial-distal width | 8.34 | 7.21 | 7.40 | 11.78 |
| Max. buccal-lingual width | 8.20 | 6.70 | 6.60 | 8.76e |
| TABLE 1. Measurements of neoceratopsian teeth from the Early-middle Cretaceous of North America. Measurements are in mm. | ||||
The enameled side of neoceratopsian tooth crowns is greatly curved in mesial or distal view (as seen in Leptoceratops, USNM 13863 and Montanoceratops, MOR 542), while in ornithopods this curvature is seen to a lesser degree (for example: Dryosaurus, GPIT 1713/21-29; Camptosaurus, UUVP 5946). The Utah and Maryland teeth exhibit curvature equal to that of the neoceratopsians.
Of primary importance is the shape of the non-enameled (or less-enameled) side of the tooth, again in lateral view. All ceratopsian teeth, including those of Psittacosauridae, are convex on this surface in both dorsoventral and mesiodistal directions (Sereno et al., 1988; Sereno and Shichin, 1988). In nearly all ornithopod teeth, this surface is concave, at least in a dorsoventral direction, and usually in an antero-posterior direction as well. The only exception is the hadrosaurid tooth shape, which is highly derived and extremely laterally compressed (Weishampel and Homer, 1990). The teeth reported here exhibit the ceratopsian condition.
The Cedar Mountain and Arundel teeth have a primary ridge that is fairly prominent, and extends to the base of the crown. ln all of the teeth it is displaced from the center, and in two (USNM 337977, MNA v9023) it is narrow and angled toward the larger of the two indentations. In the other two, the primary ridge is straight and slightly more bulbous in shape. In Tenontosaurus, the primary ridge is also prominent (OU 8-0-S3). It is less prominent, however, in other ornithopod taxa (including heterodontosaurids). Leptoceratops (USNM 13864, YPM 18133) and Udanoceratops (Kurzanov, 1982) both exhibit primary ridges that are of equal prominence to the new specimens, and that are inclined obliquely, as in USNM 337977 and MNA v9023. The primary ridge on teeth of Protoceratops and Bagaceratops is more prominent than that of the Arundel teeth (Maryanska and Osmolska, 1975). Secondary ridges are present on the Arundel teeth--three on each side of the primary ridge-that grade slowly into the indentations of the crown. This number is less on the Cedar Mountain teeth--two on each side of OMNH 28179 and one only on one side of MNA v9023. They are more prominent and shorter, as well as easier to differentiate, on USNM 337977. In USNM 497708 and the Cedar Mountain teeth, the secondary ridges extend down into the deepest part of the indentations, but still end within them, not reaching the cingulum. This trait appears to be shared by all neoceratopsian taxa. In contrast, in Tenontosaurus (OU 8-0-S3), the secondary ridges are few, and extend with equal prominence down the length of the crown. Camptosaurus (UUVP 5946) is the only ornithopod in which the secondary ridges end prior to the base of the crown, but the indentations in teeth of this taxa become shallower nearer to the root, thus causing the termination of the ridges before the base of the crown. Heterodontosaurs, Telmatosaurus, and hadrosaurs have no secondary ridges, and in Iguanodon and Dryosaurus they are very weak. Psittacosaurs have only denticles; Protoceratops, Bagaceratops, and Asiaceratops have many long secondary ridges. Leptoceratops and Montanoceratops have ridges like those of Tenontosaurus--equal prominence throughout their length--but they all end within the indentations.
In the Arundel teeth, the cingulum at the base of the crown is more prominent in USNM 497708 than in USNM 337977 (Figs.2-3). ln the former, the effect is a truncation of the primary ridge before it reaches the root, and deeper indentations. This condition is also seen in the Utah teeth, although this area is not preserved well on either specimen. Most neoceratopsian teeth have indentations that are deepest at the base of the crown (Maryanska and Osmolska, 1975). In fact, USNM 337977 is one of the few that do not exhibit this character (Bagaceratops additionally does not-Maryanska and Osmolska, 1975). Ornithopod teeth have very shallow indentations and weak cingula, if any at all.
Wear facets are present on OMNH 28179 and USNM 337977. On the former, the facet shows the typical oblique shear seen in both neoceratopsians and ornithopods. USNM 337977 exhibits a vertical shear wear pattern. This pattern is seen in ceratopsid teeth, as well as the basal neoceratopsians Montanoceratops (Chinnery and Weishampel, in press), Leptoceratops (Sternberg,1951), Udanaoceratops (Kurzanov, 1992), and also on the Zuni ceratopsian material (Wolfe et al., 1997; Chinnery et al., 1998). Wear on ornithopod teeth is never vertical it ranges from 10 to 80° from the horizontal (Weishampel, pers. comm; see also Weishampel and Witmer, 1990; Sues and Norman, 1990; Norman and Weishampel, 1990)
|
|
FIGURE 1. Neoceratopsian teeth from the Mussentuchit Member of the Cedar Mountain Formation, Utah. A-D, OMNH 28179, dentary tooth?: A) caudal view, B)lateral view, C) close up of reticulate enamel texture, D medial view; E-F, MNA v9023, maxillary tooth?: E) lateral view, F)caudal view. Scale for A, B, E, F = 1cm, for C=1 mm. |
Differences exist between the two Arundel teeth. USNM 337977 has three secondary ridges on the smaller side of the crown, and three on the larger side, with one denticle between them and several both anterior and posterior of the secondary ridges. The primary ridge is obliquely oriented, toward the larger indentation, and grades smoothly into the root. The indentations are fairly deep but stay at the same depth throughout the length of the crown. USNM 497708 has a more distinct cingulum and indentations which are deepest near the beginning of the root. There are the same number of secondary ridges, but they are more difficult to differentiate, as they extend much further into the indentations, and grade into each other. The entire enameled surface of this tooth has much more texture.
It may be that USNM 337977 is more weathered, accounting for the smaller cingulum and more shallow indentations, but the apex of the tooth and the wear facet do not exhibit signs of weathering. More probable is that the differences are due to variation among individuals or within the tooth row. Maryanska and Osmolska (1975) document a great deal of variation within basal neoceratopsian jaws depending on tooth position, and this is also seen in specimens of Leptoceratops and Bagaceratops. Overall, the two Arundel teeth resemble each other more than either resemble teeth of the other taxa studied.
The Cedar Mountain teeth are also different from the Maryland teeth and from each other. The smaller of the two, MNA v9023, exhibits neoceratopsian traits that are not as well developed as in all adult neoceratopsian teeth, leading to the conclusion that this tooth is from an immature animal or from the front or back of the tooth row (Maryanska and Osmolska, 1975). It is similar in shape to OMNH 26179, but much smaller, and is not complete. The immature nature of MNA v9023 does not allow for comparison with the other, more well developed teeth. OMNH 28179 is wider in an anteroposterior direction than the Arundel teeth. The primary ridge is wider at the base, and there are fewer secondary ridges. The indentations are also more unequal in size than in the Maryland teeth. Differences are expected, however, among teeth from such different geographic areas.
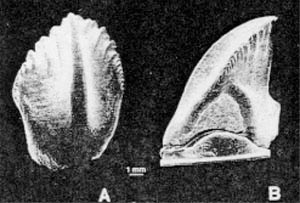 |
FIGURE 2. USNM 337977, neoceratopsian tooth from the Arundel Formation of Maryland. A) lateral view, B) caudal view. Scale = 1 mm. |
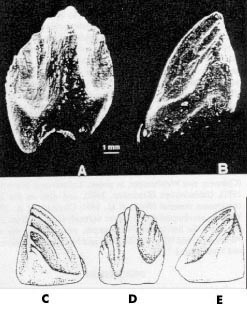 |
FIGURE 3. USNM 497708, neoceratopsian tooth from the Arundel Formation of Maryland. A, D) lateral view; B, E) caudal view; C) cranial view. Illustrations by Tracy Ford. Scale = 1 mm. |
The first Arundel tooth to be found, USNM 337977, was tentatively named by Kranz (1996). However, no more material besides the one other tooth has been identified, and the high amount of variability among neoceratopsian teeth casts doubt on the validity of the name. According to Maryanska and Osmolska (1975, p. 162): "The shape of the protoceratopsid tooth and the number and the character of the smaller, lateral ribs are variable even in the adult individuals and vary accordingly to the place which the tooth occupies in the jaw. For these reasons, we recommend that isolated protoceratopsid teeth should not be used as a basis for the erection of new taxa." Therefore, we regard the name given to the original specimen as a nomen dubium.
The biogeography of Neoceratopsia is becoming more complicated as new taxa are discovered (Chinnery et al., 1998). The clade most likely originated in Asia, as this is where the sister group of Neoceratopsia, Psittacosauridae, is distributed (Sereno, 1990). Neoceratopsian taxa are known from the Early Cretaceous of Asia (Dong and Azuma, 1997), and now from the Early Cretaceous of both eastern and western North America. Until these and other recent discoveries (Wolfe et al., 1997; Weishampel et al., in prep.), the prevailing hypothesis included a radiation of basal neoceratopsians through the late Early Cretaceous and early Late Cretaceous of Asia, and two or more dispersal events across the Bering land bridge during the middle Cretaceous (Chinnery and Weishampel, in press). These dispersal events gave rise to the derived ceratopsids known from the Late Cretaceous of western North America.
The specimens discussed in this paper necessitate a new interpretation of neoceratopsian biogeography. Three scenarios could account for the current distribution of neoceratopsian taxa. At least one of the dispersal events from Asia to North America may have occurred earlier than previously thought. This would require a route via the Bering land bridge, which has only been determined to be in place from the middle Cretaceous onward (Cifelli et al., 1997), or another land bridge in place earlier in time. Also necessary in this scenario is a route across the North American continent, to account for neoceratopsian distribution in the Maryland area. The clade may have arrived in eastern North America first, after dispersing across Europe from Asia. This is unlikely, as there is no record of ceratopsians from Europe. Only two marginocephalian taxa are known from the Early Cretaceous of Europe-Stenopelix valdensis from Germany (Sues and Galton, 1982; Dodson, 1990) and Yaverlandia bitholus, a pachycephalosaur from England (Galton, 1971). Similarities do exist between other Arundel taxa and fossils found in the Wealden Formation of Europe (Lipka, this volume) and also between the lower Cedar Mountain fauna and the Barremian of Europe (Kirkland et al.,1997), which may argue for a northern Eurasian connection with eastern North America sometime before or during the early Aptian. Many current plate tectonic reconstructions (e.g., Smith et al., 1994) for the earliest Cretaceous through the Aptian show that Europe and North America had rifted apart in the Barremian or Hauterivian and that Europe was separated from Asia by an intracontinental seaway. However, this seaway may have receded enough by the Barremian or earliest Aptian to briefly connect Europe and Asia, and possibly North America. The third possible scenario places the origin of Neoceratopsia in North America, instead of Asia, along with an early divergence of this clade and Psittacosauridae. This possibility cannot be ruled out completely, but available evidence does not support a North American origin for the clade. Complicating this problem is the depauperate state of fossil-bearing Lower Cretaceous deposits as noted by Ostrom (1970), Brinkman et al. (in press) and Kirkland et al. (1997). Further discoveries in North America will help resolve these questions.
The specimens discussed here are only a subset of recent discoveries of neoceratopsians in North America. The eventual result of these new discoveries will be a more concise understanding of the evolution and biogeography of the clade than was previously possible.
Brown, B., 1914, Leptoceratops, a new genus of Ceratopsia from the Edmonton Cretaceous of Alberta: Bulletin of the American Museum of Natural History v. 33, p. 567-580
Chinnery, B J. and Weishampel, D. B., in press, Montanoceratops cerorhynchus (Dinosauria: Ceratopsia) and relationships among basal neoceratopsians: Journal of Vertebrate Paleontology.
Chinnery, B. J., Wolfe, D. G. and Kirkland, J. I., 1998, A new consideration of neoceratopsian biogeography; in Wolberg, D. L., K. Gittis, S. Miller, L. Carey, and A. Raynor, eds., The Dinofest Symposium April 17-19, 1998 Abstracts of Presentations, p. 7.
Cifelli, R. L., Kirkland, J. I., Weil, A., Deino, A. L. and Kowallis, B. J., 1997, High-precision 40Ar/39Ar geochronology and the advent of North America's Late Cretaceous terrestrial fauna: Proceedings of the National Academy of Sciences, v. 94, p. 11163-11167.
Cifelli, R. L., Nydam, R. L., Gardner, J. D., Weil, A., Eaton, J. G., Kirkland, J. I. and Madsen, S. K., in press, Medial Cretaceous vertebrates from the Cedar Mountain Formation, Emery County; The Mussentuchit local fauna; in Gillette, D. D. ed., Vertebrate fossils of Utah: Utah Geological Survey Bulletin.
Dodson, P., 1990, Marginocephalia; in Weishampel, D. B., P. Dodson, and H. Osmolska, eds. The Dinosauria. University of California Press, p. 562-563.
Dong, Z. and Azuma, Y., 1997, On a primitive neoceratopsian from the Early Cretaceous of China; in Dong, Z. ed., Sino-Japanese Silk Road Dinosaur Expedition. China Ocean Press, Beijing, p. 68-89.
Doyle, J. A., 1992, Reversed palynological correlation of the Lower Potomac Group (USA) and the Cocobeach sequence of Gabon (Barremian-Aptian): Cretaceous Research, v. 13, p. 337-349.
Galton, P. M., 1971, A primitive dome-headed dinosaur (Ornithischia: Pachycephalosauridae) from the Lower Cretaceous of England and the function of the dome of pachycephalosaurids: Journal of Paleontology, v. 45, p. 40-46.
Gilmore, C. W., 1939, Ceratopsian dinosaurs from the Two Medicine Formation, Upper Cretaceous of Montana: Proceedings of the United States National Museum, v. 87, p. l-18.
Kirkland, J I., Britt, B., Burge, D., Carpenter, K., Cifelli, R., Decourten, F., Eaton, J., Hasiotis, S., and Lauton, T., 1997, Lower to middle Cretaceous dinosaur faunas of the central Colorado Plateau: A key to understanding 35 million years of tectonics, sedimentology; evolution and biogeography, Brigham Young University Geology Studies, v. 42, pt. II, p. 69-103.
Kranz, P. M., 1996, Notes on the sedimentary iron ores of Maryland and their dinosaurian fauna: Maryland Geological Survey Special Paper 3, p. 87-115.
Kurzanov, C. M., 1992, A gigantic protoceratopsid from the Upper Cretaceous of Mongolia: Paleontological Journal, v. 3, p. 81-93.
Maryanska, T. and Osmolska, H., 1975, Results of the Polish-Mongolian Palaeontological Expeditions-Part VI. Protoceratopsidae (Dinosauria) of Asia: Palaeontologia Polonica, v. 33, p. 133-182.
Norman, D. B., 1984, On the cranial morphology and evolution of ornithopod dinosaurs: Symposium of the Zoological Society of London, v. 52, p. 521-547.
Norman, D. B. and Weishampel, D. B., 1990, Iguanodontidae and related ornithopods; in Weishampel, D. B., P. Dodson, and H. Osmolska, eds., The Dinosauria. University of California Press, p. 510-533.
Obradovich, J , 1993, A Cretaceous time scale; in Caldwell, W. G. E., and E. G. Kauffman, eds., Evolution of the Western Interior Basin, Geological Association of Canada Special Paper 39, p. 379-396.
Ostrom, J. H., 1970, Stratigraphy and paleontology of the Cleverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana, Yale Peabody Museum Bulletin 35, 234 pp.
Sereno, P. C., 1990, Psittacosauridae; in Weishampel, D. B., P. Dodson, and H. Osmolska, eds., The Dinosauria. University of California Press, p. 579-592.
Sereno, P. C., 1986, Phylogeny of the bud-hipped dinosaurs (Order Ornithischia). National Geographic Research v. 2, p 234-256.
Sereno, P. C. and Shichin, C., 1988, Psittacosaurus meileyingensis (Ornithischia: Ceratopsia), a new psittacosaur from the lower Cretaceous of northwestern China: Journal of Vertebrate Paleontology, v. 8, p. 353-365.
Sereno, P. C., Shichin, C., Zhengwu, C., and Chenggang, R., 1988, Psittacosaurus meileyingensis (Ornithischia: Ceratopsia), a new psittacosaur from the Lower Cretaceous of northeastern China, Journal of Vertebrate Paleontology, v. 8, p. 366-377.
Smith, A. G., Smith, D. G., and Funnell, B. M., 1994, Atlas of Mesozoic and Cenozoic coastlines Cambridge University Press, 99 pp.
Sternberg, C. M., 1951, Complete skeleton of Leptoceratops gracilis Brown from the upper Edmonton Member on Red Deer River, Alberta: Bulletin of the National Museum of Canada, v. 123, p. 225-255.
Stokes, W. L., 1952, Lower Cretaceous in Colorado Plateau: American Association of Petroleum Geologists Bulletin 36, p. 1766-1776.
Sues, H.-D. and Galton, P. M., 1982, The systematic position of Stenopelix valdensis (Reptilia: Ornithischia) from the Wealden of north-western Germany: Palaeontographica, v. 178, p. 183-190.
Sues, H.-D. and Norman, D. B., 1990, Hypsilophodontidae, Tenontasaurus, Dryosauridae; in Weishampel, D. B., P. Dodson, and H. Osmolska, eds., The Dinosauria, University of California Press, p. 498-509.
Weishampel, D. B., 1990, Ornithopoda; in Weishampel, D. B., P. Dodson, and H. Osmolska, eds., The Dinosauria. University of California Press, p. 484-485.
Weishampel, D. B. and Horner, J. R., 1990, Hadrosauridae; in Weishampel, D. B., P. Dodson, and H. Osmolska, eds., The Dinosauria. University of California Press, p. 534-561.
Weishampel, D. B., Meers, M. B., Akersten, W. A. and McCrady, A. D., in preparation, New Early Cretaceous dinosaur remains, including possible ceratopsians, from the Wayan Formation of eastern Idaho.
Weishampel, D. B., Norman, D. B. and Grigorescu, D., 1993, Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: The most basal hadrosaurid dinosaur: Paleontology, v. 36, p. 361-385.
Weishampel, D. B. and Witmer, L. M., 1990, Heterodontosauridae; in Weishampel, D. B., P. Dodson, and H. Osmolska, eds., The Dinosauria. University of California Press, p. 486-497.
Wolfe, D. G., Kirkland, J. I., Denton, R. and Anderson, B. G., 1997, A new terrestrial vertebrate record from the Moreno Hill Formation (Turonian, Cretaceous), west-central New Mexico: Journal of Vertebrate Paleontology, v. l7 suppl, p. 85-86A.
Young, R. G., 1960, Dakota Group of the Colorado Plateau: American Association of Petroleum Geologists Bulletin 44, p. 156-194.